Адаптация микроорганизмов к условиям внешней среды. Бактерии
ПРИКЛАДНАЯ БИОХИМИЯ И МИКРОБИОЛОГИЯ, 2004, том 40, № 4, с. 387-397
УДК: 576.8.098/577.1
ВНЕКЛЕТОЧНЫЕ ФАКТОРЫ АДАПТАЦИИ БАКТЕРИЙ К НЕБЛАГОПРИЯТНЫМ УСЛОВИЯМ СРЕДЫ
© 2004 г. Ю. А. Николаев
Институт микробиологии РАН, 117811, Москва, e-mail: [email protected] Поступила в редакцию 17.11.2003 г.
Рассмотрены сведения о внеклеточных соединениях бактерий, участвующих в их адаптации к неблагоприятным условиям среды: высоким и низким температурам, рост-ингибирующим и бактерицидным концентрациям токсичных веществ (окислители, фенолы, тяжелые металлы), антибиотиков, неблагоприятным значениям рН и солености. Идентифицированные соединения по химической природе относятся к разным типам, они представлены белками, углеводородами, органическими кислотами, нуклеотидами, аминокислотами, липопептидами, летучими соединениями. Большая часть таких соединений в настоящее время не идентифицирована, а свойства их изучены при использовании биотестов. Предлагается рассматривать внеклеточные факторы адаптации (ВФА), как новую группу биологически активных веществ. По механизму действия внеклеточные факторы адаптации можно разделить на несколько групп; протекторы (стабилизаторы); вещества сигнальной природы, являющиеся индукторами защитных механизмов клетки; регуляторы - не индукторы, (например, регуляторы адгезии); "противоядия" и нейтрализующего действия. Основными направлениями изучения ВФА являются поиск новых соединений (на основе биотестов), их идентификация и исследование механизмов действия. Внеклеточные факторы адаптации могут найти широкое практическое применение в биотехнологии, медицине, сельском хозяйстве и охране окружающей среды.
Адаптация бактерий к неблагоприятным условиям среды является традиционным и хорошо изученным разделом биохимии и микробиологии. Под адаптацией (лат. айаргагю - приспособление) понимают сумму физиологических, биохимических, морфологических и поведенческих реакций организма, направленных на изменение скорости роста, метаболизма, жизнеспособности (выживаемости) и генетически присущих организму. Адаптация направлена на выживание конкретной популяции и всего вида в целом. В учебниках по микробиологии, биохимии и теоретической биологии адаптация к неблагоприятным условиям среды рассматривается в разделах "фенотипичес-кая и генетическая адаптации" и "регуляция активности ферментов и их синтеза". Конкретные примеры адаптации многообразны и описаны в ряде обзоров и монографий, в основном с позиций биохимии и генетического контроля развития адаптивного ответа .
Кратко остановимся на терминологии, т.к. даже среди специалистов, работающих в этой области, отсутствует единство в употреблении конкретных терминов. В англоязычной литературе обычно говорят об адаптации как о развитии устойчивости к стрессам или шокам (кислотному, температурному, солевому и др.), понимая под стрессом давление, напряжение, нажим, другими словами - существенное изменение какого-то фактора - температуры, давления и т.п. "Шок"
означает удар, потрясение, толчок, т.е. резкое воздействие на организм, кратковременное по сравнению с длиной клеточного цикла и скоростью обычных адаптивных реакций, и значительное по интенсивности воздействия фактора.
В отечественной биологии существует иная терминология. Согласно Большому толковому словарю русского языка "стресс - это состояние напряжения организма, защитная реакция, вызванная действием неблагоприятного фактора". Такое понимание согласуется с определением основоположника стрессологии Г. Селье, который трактовал стресс как неспецифический ответ организма на предъявленное ему требование . В таком же значении стресс рассматривается в ряде обзоров . Шок - это "реакция организма на сильное внешнее воздействие (а также его состояние после такого воздействия), характеризующаяся резким нарушением регуляции жизненных процессов" . Таким образом, и стресс и шок - это состояние организма, характерная адаптивная реакция на воздействия, различающиеся дозой, интенсивностью и временем.
Правомочно коснуться вопроса о том, насколько состояние стресса является естественным, распространенным или анормальным. В таком случае следует обозначить, что принимается за норму. Постоянное изменение, развитие - неотъемлемые свойства биологических систем .
Изменения носят как векторный, однонаправленный характер, так и циклический. Соответственно, в развитии любой живой системы должны чередоваться фазы быстрого, оптимального развития и фазы подавленного, неоптимального. Исходя из этого, подавленные, лимитированные состояния следует считать естественным и даже неотъемлемым свойством жизни . В таком случае, если принять за "норму" размножение микроорганизмов, увеличение их численности в фазе роста микробной культуры, то состояние клеток в лаг-фазе рассматривается как "стресс новой среды", требующий адаптивных приспособительных реакций. С другой стороны, истощение источников питания или критическое повышение плотности клеток в развивающейся культуре обусловливают "стресс голода", а стационарные клетки демонстрируют физиологическое приспособление культуры к неоптимальным для роста условиям. Такое рассмотрение стресса как нормального и даже способствующего развитию состояния культуры (организма) обусловлено и сочетается с циклическим характером того конкретного физического мира, в котором существуют земные организмы, где циклически изменяются температура, освещенность, влажность, давление, концентрации органических и неорганических соединений, напряженность физических полей, действие биотических факторов.
Принимая во внимание цикличность изменений окружающих условий, следует выделять в них: а) изменения, новые для конкретного этапа развития, но повторяющиеся в цикле развития и входящие в зону толерантности данного вида; б) выходящие за пределы такой зоны толерантности воздействия, неблагоприятные для роста и развития организма, часто биоцидные. Тогда, под адаптацией к каким-либо условиям будем понимать совокупность специфических морфологических, биохимических, физиологических и поведенческих реакций организма, развивающихся в ответ на данные условия и способствующих продолжению функционирования организма или направленные на повышение жизнеспособности (снижение смертности) в экстремальных условиях. В последнем случае речь идет не о продолжении метаболизма (микробной культуры) в конкретных неблагоприятных условиях (адаптации организма), а о сохранении популяции для следующего жизненного цикла в отдаленном будущем, при гибели части популяции (как правило, большей) и временной остановке в функционировании переживающих клеток.
В контексте обсуждаемой проблемы кратко остановимся на признаках стресса у микроорганизмов. При его констатации обязательно сравнение показателей состояния организма, которые наблюдаются в обычном, принимаемом за оптимальное (гомеостаз), и в стрессовом состоя-
нии. Значительное нарушение любого измеряемого показателя гомеостаза является показателем и признаком стресса. У микроорганизмов к показателям угнетенного, стрессового состояния относят: выход белка во внеклеточное пространство , потерю низкомолекулярных соединений клеткой вследствие повышения проницаемости цитоплазматической мембраны, повреждение рибосом, нуклеиновых кислот, уменьшение скорости потребления кислорода, снижение активности ферментов , накопление активных форм кислорода и продуктов перекисного окисления липидов , потерю частью клеток популяции способности образовывать колонии при росте на плотных питательных минимальных средах (т.е. снижение концентрации ко-лониеобразующих единиц, КОЕ) , замедление роста , торможение жизненной активности , агрегацию и адгезию. Обязательным признаком стрессового состояния является его обратимость, возможность возврата к нормальному функционированию при соответствующих изменениях среды.
Среди перечисленных признаков наиболее характерные и чаще всего используемые на практике два - снижение скорости роста и жизнеспособности клеток. Они представляются наиболее прямыми и адекватными показателями состояния стресса. Скорость роста - это интегральный показатель состояния микроорганизмов. Для клеток, растущих с определенной, максимальной для данных условий скоростью (ц.макс), ее снижение до какой-то более низкой величины цмин будет свидетельствовать о стрессовом состоянии микробов. Однако впоследствии рост с этой новой низкой скоростью может стать нормой в новых условиях. Дальнейшее ухудшение условий может привести к снижению ц до 0 или даже к гибели клеток. Начало гибели клеток свидетельствует об исчерпании адаптивных ресурсов отдельных клеток. Однако для популяции уменьшение количества клеток - процесс обратимый и гибель определенного количества отдельных клеток -вполне нормальное явление при том, что другая часть клеток сохраняет жизнеспособность, переходя в покоящееся состояние. Таким образом, снижение скорости роста и концентрации жизнеспособных клеток являются признаками стресса, но первый более характеризует состояние клеток, а второй - популяции. Большинство работ, посвященных адаптации бактерий к неблагоприятным воздействиям, оперирует именно этими двумя показателями.
Обзор ограничен, в основном, адаптацией активно растущих культур к неблагоприятным физико-химическим условиям среды, направленной как на поддержание активного состояния клеток, когда стратегия роста сохраняется, так и на сохранение популяции при смене стратегии роста
стратегией переживания. Адаптации микробных культур к таким условиям, как истощение питательных веществ, новые благоприятные условия, смена источников питания, часто сопряженным с онтогенезом культур (т.е. образованием и прорастанием покоящихся форм) не будут рассмотрены.
В настоящее время достаточно хорошо исследованы механизмы адаптации бактерий к высокой и низкой температурам, высоким концентрациям активных форм кислорода, солей, неионных веществ, высокой радиации, гидростатическому давлению , исчерпанию источников углерода, энергии и других ресурсов . Основное внимание уделяется внутриклеточным из
ОЛЕСКИН А.В. - 2009 г.
ЗАЙЦЕВА Ю.В., ПОПОВА А.А., ХМЕЛЬ И.А. - 2014 г.

 Реакция на стрессовые воздействия В процессе эволюции бактерии, так же как и любые другие живые организмы, приспособились к существованию в условиях не вполне оптимальных, а иногда и таящих опасность для жизни. Токсические вещества, неблагоприятная температура, р. Н, облучение в пределах, определяемых видовой или штаммовой чувствительностью организма, не препятствуют нормальному существованию бактерий. Резкие изменения условий в неблаго приятную торону с приводят к отмиранию клеток. Однако при некоторых воздействиях, которые обычно обозначают как сублетальные, клетки не погибают сразу, а оказываются травмированными. Их дальнейшая судьба в значительной степени зависит от условий, в которые они окажутся.
Реакция на стрессовые воздействия В процессе эволюции бактерии, так же как и любые другие живые организмы, приспособились к существованию в условиях не вполне оптимальных, а иногда и таящих опасность для жизни. Токсические вещества, неблагоприятная температура, р. Н, облучение в пределах, определяемых видовой или штаммовой чувствительностью организма, не препятствуют нормальному существованию бактерий. Резкие изменения условий в неблаго приятную торону с приводят к отмиранию клеток. Однако при некоторых воздействиях, которые обычно обозначают как сублетальные, клетки не погибают сразу, а оказываются травмированными. Их дальнейшая судьба в значительной степени зависит от условий, в которые они окажутся.
 Травмирование клеток происходит под влиянием повышенной или понижен ной температуры, под действием токсических веществ в не слишком высоких концентрациях, в результате голодания, при осмотическом шоке, облучениях. У травмированных клеток во многих случаях нарушаются барьерные функции мембран, наблюдается выход в среду некоторых метаболитов, нарушается синтез белка, возникают нарушения в структуре ДНК. Некото рые условия, вполне благоприятные для развития нормальных бактерий. могут быть
Травмирование клеток происходит под влиянием повышенной или понижен ной температуры, под действием токсических веществ в не слишком высоких концентрациях, в результате голодания, при осмотическом шоке, облучениях. У травмированных клеток во многих случаях нарушаются барьерные функции мембран, наблюдается выход в среду некоторых метаболитов, нарушается синтез белка, возникают нарушения в структуре ДНК. Некото рые условия, вполне благоприятные для развития нормальных бактерий. могут быть
 Например, бактерии, подвергнутые сублетальному температурному шоку, осмотичсскому шоку и других воздействий, гибнут на средах с повышенной концентрацией солей, со вершенно не опасной для нормальных клеток, или на средах с поверхностно активными соединениями, также в концентрациях, не влияющих на рост нормальных клеток. Эти факторы иногда определяют как селектирующие здоровые клетки от травмиро ванных. Травмированные клетки, помещенные в благоприятные ус ловия, способны епарировать, р
Например, бактерии, подвергнутые сублетальному температурному шоку, осмотичсскому шоку и других воздействий, гибнут на средах с повышенной концентрацией солей, со вершенно не опасной для нормальных клеток, или на средах с поверхностно активными соединениями, также в концентрациях, не влияющих на рост нормальных клеток. Эти факторы иногда определяют как селектирующие здоровые клетки от травмиро ванных. Травмированные клетки, помещенные в благоприятные ус ловия, способны епарировать, р
 Прямая фотореактивация наблюдается при освещении клеток УФ светом, при этом происходит разрезании пиримидиновых димеров в ДНК, вот поэтому осо бое начение з для фотореактивации имеет УФ. Процесс фотореактивации связан с действием фермента фотолиазы, являющейся флавопротеином. Фотолиаза связывается с пиримидиновыми димерами, активация фермент субстратного комплекса светом длиной волны 300 - 600 нм приводит к мономеризации димеров. При этом корреляции между общей радиорезистентностью клетки и способностью к
Прямая фотореактивация наблюдается при освещении клеток УФ светом, при этом происходит разрезании пиримидиновых димеров в ДНК, вот поэтому осо бое начение з для фотореактивации имеет УФ. Процесс фотореактивации связан с действием фермента фотолиазы, являющейся флавопротеином. Фотолиаза связывается с пиримидиновыми димерами, активация фермент субстратного комплекса светом длиной волны 300 - 600 нм приводит к мономеризации димеров. При этом корреляции между общей радиорезистентностью клетки и способностью к
 Наряду с описанной прямой имеет место непрямая фотореактивация с пиком в области 340 нм, который не оказывает влияния на расщепление димеров, а происходит задержка роста бактерий, в результате чего удлиняется период протекания репарационных процессов. Травмированные клетки не просто восстанавливают причиненные им повреждения, также под влиянием сублетальных воздействий неблагоприятных факторов производят перестройку в обмене веществ Клетки, под вергшиеся неблагоприятным воздействиям, находятся в состоянии стресса. В различных случаях при стресссе может быть связано или не связано с нарушениями клеточных структур, т. е. клетки могут быть или не быть травмированы.
Наряду с описанной прямой имеет место непрямая фотореактивация с пиком в области 340 нм, который не оказывает влияния на расщепление димеров, а происходит задержка роста бактерий, в результате чего удлиняется период протекания репарационных процессов. Травмированные клетки не просто восстанавливают причиненные им повреждения, также под влиянием сублетальных воздействий неблагоприятных факторов производят перестройку в обмене веществ Клетки, под вергшиеся неблагоприятным воздействиям, находятся в состоянии стресса. В различных случаях при стресссе может быть связано или не связано с нарушениями клеточных структур, т. е. клетки могут быть или не быть травмированы.
 Воздейст вия, которые приводят летки в к состояние стресса, определяют как стрессорные. Процессы, протекающие в клетках, находящихся в состоянии стресса, изучены преимущественно на модели кишечных бактерии, прежде всего Е. coli и Salmonella. К на стоящему времени у кишечных бактерий выявлено 5 регуляторных систем ответа на стрессовые воздействия: «строгий контроль» ; SOS ответ; адаптивный ответ;
Воздейст вия, которые приводят летки в к состояние стресса, определяют как стрессорные. Процессы, протекающие в клетках, находящихся в состоянии стресса, изучены преимущественно на модели кишечных бактерии, прежде всего Е. coli и Salmonella. К на стоящему времени у кишечных бактерий выявлено 5 регуляторных систем ответа на стрессовые воздействия: «строгий контроль» ; SOS ответ; адаптивный ответ;
 синтез белков теплового шока; ответ на окислительный стресс. Во всех 5 ти перечисленных случаях происходят глубокие перестройки мета болизма, связанные с замедлением или прекращением размно жения и синтезом белков, необходимых для выживания. В некоторых случаях, в процессах регуляции, при нимают участие специальные соединения, клеточные гормоны, получившие название алармонов (фр. alarme-тревога).
синтез белков теплового шока; ответ на окислительный стресс. Во всех 5 ти перечисленных случаях происходят глубокие перестройки мета болизма, связанные с замедлением или прекращением размно жения и синтезом белков, необходимых для выживания. В некоторых случаях, в процессах регуляции, при нимают участие специальные соединения, клеточные гормоны, получившие название алармонов (фр. alarme-тревога).
 Адаптация или абаптация Естественный отбор приводит к соответствию организмы и среду их обитания. Исход взаимодействия между организмом и средой может быть различным: от широкого значения до исчезновения вида. В одних условиях вид выживает и размножается, в других – нет. В этом смысле природа осуществляет отбор. Не лишено смысла утверждение, что особи данного поколения «абаптированы» условиям, в которых жили предыдущие поколения. Условия прошлого – фильтр, через который некоторые сочетания признаков просочились в настоящее.
Адаптация или абаптация Естественный отбор приводит к соответствию организмы и среду их обитания. Исход взаимодействия между организмом и средой может быть различным: от широкого значения до исчезновения вида. В одних условиях вид выживает и размножается, в других – нет. В этом смысле природа осуществляет отбор. Не лишено смысла утверждение, что особи данного поколения «абаптированы» условиям, в которых жили предыдущие поколения. Условия прошлого – фильтр, через который некоторые сочетания признаков просочились в настоящее.
 Термин «приспособление» («адаптация») оставляет ложное впечатление какого то предсказания, предвидения или, по крайней мере, замысла. Организмы не предназначены, не адаптированы ни для настоящего, ни для будущего и ни настоящему, ни к будущему не приспособлены – они являют собой живые следствия собственного прошлого. Они абаптированы своим прошлым. Приспособленность – это относительный выпад особей в численность будущих поколений. Наиболее приспособленные особи популяции те, численность потомства которых наиболее велика по сравнению с численностью потомства,
Термин «приспособление» («адаптация») оставляет ложное впечатление какого то предсказания, предвидения или, по крайней мере, замысла. Организмы не предназначены, не адаптированы ни для настоящего, ни для будущего и ни настоящему, ни к будущему не приспособлены – они являют собой живые следствия собственного прошлого. Они абаптированы своим прошлым. Приспособленность – это относительный выпад особей в численность будущих поколений. Наиболее приспособленные особи популяции те, численность потомства которых наиболее велика по сравнению с численностью потомства,
 Адаптация микроорганизмов - изменение строения и физиологии микробов под действием факторов окружающей среды. Степень адаптации микроорганизма к новым условиям зависит от стойкости данной культуры и силы действия внешних факторов окружающей среды. Некоторые виды микробов, например кислото спирто щёлочеустойчивые, дифтерийная группа и грибки изменяются несколько меньше, тогда как тифозная группа, кокки и анаэробы группы легче приспосабливаются.
Адаптация микроорганизмов - изменение строения и физиологии микробов под действием факторов окружающей среды. Степень адаптации микроорганизма к новым условиям зависит от стойкости данной культуры и силы действия внешних факторов окружающей среды. Некоторые виды микробов, например кислото спирто щёлочеустойчивые, дифтерийная группа и грибки изменяются несколько меньше, тогда как тифозная группа, кокки и анаэробы группы легче приспосабливаются.
 Приспособляемость микробов в наибольшей степени выражается в отношение температуры и наличию кислорода, адаптация идёт тем лучше и совершенее, чем медленее и постепеннее нарастает воздействие новых факторов. Новые условия могут заставить микроорганизмы стать менее требовательным к среде, ограничивать свои физиологические потребности или образовывать споры, иногда изменяется морфология микробной клетки и её строение.
Приспособляемость микробов в наибольшей степени выражается в отношение температуры и наличию кислорода, адаптация идёт тем лучше и совершенее, чем медленее и постепеннее нарастает воздействие новых факторов. Новые условия могут заставить микроорганизмы стать менее требовательным к среде, ограничивать свои физиологические потребности или образовывать споры, иногда изменяется морфология микробной клетки и её строение.
 Возникновение микробных штаммов обладающих новыми свойствами происходит особенно часто при столкновении с вредными химическими соединениями в среде, в которой микроорганизмы обычно свободно размножаются - при этом часть из них погибает, а наиболее стойкие выживают и дают стойкие или резистентные штаммы, иногда даже по отношению к антибиотикам и иммунным сывороткам.
Возникновение микробных штаммов обладающих новыми свойствами происходит особенно часто при столкновении с вредными химическими соединениями в среде, в которой микроорганизмы обычно свободно размножаются - при этом часть из них погибает, а наиболее стойкие выживают и дают стойкие или резистентные штаммы, иногда даже по отношению к антибиотикам и иммунным сывороткам.
 Вывод: адаптация это: 1) эволюционно возникшее приспособление организма к условиям среды, выражающееся в изменении их внешних и внутренних особенностей (в биологии); 2) любое приспособление органа, функции или организма к изменяющимся условиям среды (в медицине); 3) совокупность реакций живой системы, поддерживающих ее функциональную устойчивость при измерении условий среды, окружающих эту систему.
Вывод: адаптация это: 1) эволюционно возникшее приспособление организма к условиям среды, выражающееся в изменении их внешних и внутренних особенностей (в биологии); 2) любое приспособление органа, функции или организма к изменяющимся условиям среды (в медицине); 3) совокупность реакций живой системы, поддерживающих ее функциональную устойчивость при измерении условий среды, окружающих эту систему.
 Хемотаксис бактерий Подвижные бактерии активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие направленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис. Наибольшее внимание привлекает изучение хемотаксиса, т. е. движения в определенном направлении относительно источника химического вещества. Для каждого организма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы (эффекторы). Среди последних выделяют аттрактанты (вещества, привлекающие
Хемотаксис бактерий Подвижные бактерии активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие направленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис. Наибольшее внимание привлекает изучение хемотаксиса, т. е. движения в определенном направлении относительно источника химического вещества. Для каждого организма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы (эффекторы). Среди последних выделяют аттрактанты (вещества, привлекающие
 Аттрактантами могут быть сахара, аминокислоты, витамины, нуклеотиды и другие химические молекулы; репеллентами некоторые аминокислоты, спирты, фенолы, неорганические ионы. Аттрактантом для аэробных прокариот и репеллентом для анаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны. За чувствительность бактерий к градиентам определенных факторов ответственны специфические рецепторы.
Аттрактантами могут быть сахара, аминокислоты, витамины, нуклеотиды и другие химические молекулы; репеллентами некоторые аминокислоты, спирты, фенолы, неорганические ионы. Аттрактантом для аэробных прокариот и репеллентом для анаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны. За чувствительность бактерий к градиентам определенных факторов ответственны специфические рецепторы.
 Адгезия микроорганизмов Адгезия способность микроорганизмов адсорбироваться на твердых поверхностях и чувствительных клетках с последующей колонизацией, т. е. пусковой механизм инфекционного процесса. Белковые структуры, ответственные за связывание микроорганизма с клеткой, расположены на его поверхности и называются адгезинами. Адгезины разнообразны по строению и обусловливают высокую специфичность, которая проявляется в способности одних микроорганизмов прикрепляться к клеткам эпителия дыхательных
Адгезия микроорганизмов Адгезия способность микроорганизмов адсорбироваться на твердых поверхностях и чувствительных клетках с последующей колонизацией, т. е. пусковой механизм инфекционного процесса. Белковые структуры, ответственные за связывание микроорганизма с клеткой, расположены на его поверхности и называются адгезинами. Адгезины разнообразны по строению и обусловливают высокую специфичность, которая проявляется в способности одних микроорганизмов прикрепляться к клеткам эпителия дыхательных
 На процесс адгезии могут влиять физико химические механизмы, связанные с гидрофобностью микробных клеток, суммой энергии притяжения и отталкивания. У грамотрицательных бактерий адгезия происходит за счет пилей I го и общего типов. У грамположительных бактерий адгезины представляют собой белки и тейхоевые кислоты клеточной стенки. У других микроорганизмов эту функцию выполняют различные структуры клеточной системы: поверхностные белки, липополисахариды и др. Адгезия к поверхности различных небиологических материалов обусловлена как их физико химическими свойствами, так и специфическими поверхностными рецепторами. Антиангезионная терапия, основанная на борьбе с адгезией микробов, заключается в использовании агентов,
На процесс адгезии могут влиять физико химические механизмы, связанные с гидрофобностью микробных клеток, суммой энергии притяжения и отталкивания. У грамотрицательных бактерий адгезия происходит за счет пилей I го и общего типов. У грамположительных бактерий адгезины представляют собой белки и тейхоевые кислоты клеточной стенки. У других микроорганизмов эту функцию выполняют различные структуры клеточной системы: поверхностные белки, липополисахариды и др. Адгезия к поверхности различных небиологических материалов обусловлена как их физико химическими свойствами, так и специфическими поверхностными рецепторами. Антиангезионная терапия, основанная на борьбе с адгезией микробов, заключается в использовании агентов,
 В частности, у бактерий широко распростране на так называемая горизонтальная передача генетических де терминант, при которой некоторые гены могут быть перенесены не только от одних членов популяции данного вида другим, но и представителям разных видов и даже родов. Горизонтальной передаче подвержены главным образом плазмидные гены, опре деляющие адаптации бактерий к некоторым факторам среды. Это гены устойчивости к токсическим веществам, гены патогенности, гены, определяющие способность к использованию неко торых органических соединений и водорода, и др.
В частности, у бактерий широко распростране на так называемая горизонтальная передача генетических де терминант, при которой некоторые гены могут быть перенесены не только от одних членов популяции данного вида другим, но и представителям разных видов и даже родов. Горизонтальной передаче подвержены главным образом плазмидные гены, опре деляющие адаптации бактерий к некоторым факторам среды. Это гены устойчивости к токсическим веществам, гены патогенности, гены, определяющие способность к использованию неко торых органических соединений и водорода, и др.
 Таким обра зом, генофонд популяции микробов обогащается за счет некоторых генов, имеющихся в популяции других видов. Несмотря на относительную про стоту организации бактериальной клетки и ее незначительный объем, она обладает весьма сложными и совершенными механизмами в виде молекулярных адаптаций.
Таким обра зом, генофонд популяции микробов обогащается за счет некоторых генов, имеющихся в популяции других видов. Несмотря на относительную про стоту организации бактериальной клетки и ее незначительный объем, она обладает весьма сложными и совершенными механизмами в виде молекулярных адаптаций.
 Некультивируемые формы патогенных бактерий У многих видов грамотрицательных бактерий, в том числе у патогенных (шигеллы, сальмонеллы, холерный вибрион и др.) существует особое приспособительное, генетически регулируемое состояние, физиологически эквивалентное цистам, в которое они могут переходить под влиянием неблагоприятных условий и сохранять жизнеспособность до нескольких лет. Симбиоз нескольких видов бактерий, используемых в медикаментах, хорошо помогает при лечении заболеваний.
Некультивируемые формы патогенных бактерий У многих видов грамотрицательных бактерий, в том числе у патогенных (шигеллы, сальмонеллы, холерный вибрион и др.) существует особое приспособительное, генетически регулируемое состояние, физиологически эквивалентное цистам, в которое они могут переходить под влиянием неблагоприятных условий и сохранять жизнеспособность до нескольких лет. Симбиоз нескольких видов бактерий, используемых в медикаментах, хорошо помогает при лечении заболеваний.
 Главная особенность этого состояния заключается в том, что такие бактерии не размножаются и поэтому не образуют колоний на плотной питательной среде. Такие не размножающиеся, но жизнеспособные клетки получили название некультивируемых форм бактерий (НФБ), они обладают активными метаболическими системами, в том числе системами переноса электронов, биосинтеза белка и нуклеиновых кислот, и сохраняют вирулентность. Их клеточная мембрана более вязкая, клетки обычно приобретают форму кокков, имеют значительно уменьшенные размеры.
Главная особенность этого состояния заключается в том, что такие бактерии не размножаются и поэтому не образуют колоний на плотной питательной среде. Такие не размножающиеся, но жизнеспособные клетки получили название некультивируемых форм бактерий (НФБ), они обладают активными метаболическими системами, в том числе системами переноса электронов, биосинтеза белка и нуклеиновых кислот, и сохраняют вирулентность. Их клеточная мембрана более вязкая, клетки обычно приобретают форму кокков, имеют значительно уменьшенные размеры.
 Они обладают более высокой устойчивостью во внешней среде и поэтому могут переживать в ней длительное время (например, холерный вибрион в грязном водоеме), поддерживая эндемическое состояние данного региона (водоема). Для обнаружения НФБ используют молекулярно генетические методы (ДНК-ДНК гибридизация, ЦПР), а также более простой метод прямого подсчета жизнеспособных клеток. С этой целью к исследуемому материалу добавляют в небольшом количестве питательные вещества (дрожжевой экстракт) и налидиксовую кислоту (для подавления синтеза ДНК) на несколько часов.
Они обладают более высокой устойчивостью во внешней среде и поэтому могут переживать в ней длительное время (например, холерный вибрион в грязном водоеме), поддерживая эндемическое состояние данного региона (водоема). Для обнаружения НФБ используют молекулярно генетические методы (ДНК-ДНК гибридизация, ЦПР), а также более простой метод прямого подсчета жизнеспособных клеток. С этой целью к исследуемому материалу добавляют в небольшом количестве питательные вещества (дрожжевой экстракт) и налидиксовую кислоту (для подавления синтеза ДНК) на несколько часов.
 Клетки усваивают питательные вещества и увеличиваются в размерах, но не делятся, поэтому такие увеличенные клетки четко видны в микроскоп и их легко подсчитать. Для этих целей можно использовать также методы цитохимические (образование формазана) или микроауторадиографии.
Клетки усваивают питательные вещества и увеличиваются в размерах, но не делятся, поэтому такие увеличенные клетки четко видны в микроскоп и их легко подсчитать. Для этих целей можно использовать также методы цитохимические (образование формазана) или микроауторадиографии.
 Гомеостаз и механизмы гомеостаза Гомеостаз это способность сохранять устойчивость и постоянство внутренней среды организма В основе гомеостаза лежит тонкий баланс внутри системы, при нарушении которого организм пытается найти резервы для восстановления. При нарушении равновесия система или отдельный организм рискует прекратить свое существование, поэтому должны уметь хорошо адаптироваться к внешним условиям среды и постоянно развиваться.
Гомеостаз и механизмы гомеостаза Гомеостаз это способность сохранять устойчивость и постоянство внутренней среды организма В основе гомеостаза лежит тонкий баланс внутри системы, при нарушении которого организм пытается найти резервы для восстановления. При нарушении равновесия система или отдельный организм рискует прекратить свое существование, поэтому должны уметь хорошо адаптироваться к внешним условиям среды и постоянно развиваться.
 Гомеостатические системы обладают такими свойствами как нестабильность (выбор способа для приспособления к окружающей среде), тяготение к равновесию (стремление всеми силами сохранить равновесие системы) и непредсказуемость (результат может оказаться иным, чем ожидалось). У человека за поддержание гомеостаза отвечают вегетативная нервная система и эндокринная система, которая контролируется гипоталамусом, а последний, в свою очередь, корой головного мозга.
Гомеостатические системы обладают такими свойствами как нестабильность (выбор способа для приспособления к окружающей среде), тяготение к равновесию (стремление всеми силами сохранить равновесие системы) и непредсказуемость (результат может оказаться иным, чем ожидалось). У человека за поддержание гомеостаза отвечают вегетативная нервная система и эндокринная система, которая контролируется гипоталамусом, а последний, в свою очередь, корой головного мозга.
 Существует два механизма гомеостаза, отрицательная обратная связь и положительная обратная связь. Первая меняет реакцию системы на противоположную, вторая приводит к дестабилизирующему эффекту. Пример: сохранение постоянной температуры тела, сохранение определенного числа особей внутри сообщества, повышение и понижение кровяного давления при недостатке кислорода. У сложных организмов есть ряд детекторов, способных заранее предупредить об опасной ситуации. Например, исчезновение птиц и животных с места, где вскоре должна произойти
Существует два механизма гомеостаза, отрицательная обратная связь и положительная обратная связь. Первая меняет реакцию системы на противоположную, вторая приводит к дестабилизирующему эффекту. Пример: сохранение постоянной температуры тела, сохранение определенного числа особей внутри сообщества, повышение и понижение кровяного давления при недостатке кислорода. У сложных организмов есть ряд детекторов, способных заранее предупредить об опасной ситуации. Например, исчезновение птиц и животных с места, где вскоре должна произойти
 фаза эпидемического распро странения характеризуется наиболее широким поражени ем особей в популяции хозяев. И одновременно ней происходит накопление устойчи вых к данной болезни особей - не заболевших или уже ставших иммунными в результате перенесенной болезни. стадия резервационного преобразования, где вирулентные свойства становятся не столь выгодными, поскольку активные, патогенетические механиз мы вызывают столь же активный ответ со стороны иммунной системы устойчивых к болезни хозяев. Снова включается на правленный отбор, но уже в сторону накопления
фаза эпидемического распро странения характеризуется наиболее широким поражени ем особей в популяции хозяев. И одновременно ней происходит накопление устойчи вых к данной болезни особей - не заболевших или уже ставших иммунными в результате перенесенной болезни. стадия резервационного преобразования, где вирулентные свойства становятся не столь выгодными, поскольку активные, патогенетические механиз мы вызывают столь же активный ответ со стороны иммунной системы устойчивых к болезни хозяев. Снова включается на правленный отбор, но уже в сторону накопления
 Регуляция численности патогенных микроорганизмов Можно говорить о двух типах регуляции численности патогенных микроорганизмов: 1. естественной (независимой от деятельности человека) 2. антропогенной (вызванной деятельностью человека), которые в современных условиях нередко взаимосвязаны. 3. Внутрипопуляционная регуляция: Регуляция организмом хозяина. Регуляция популяцией хозяина.
Регуляция численности патогенных микроорганизмов Можно говорить о двух типах регуляции численности патогенных микроорганизмов: 1. естественной (независимой от деятельности человека) 2. антропогенной (вызванной деятельностью человека), которые в современных условиях нередко взаимосвязаны. 3. Внутрипопуляционная регуляция: Регуляция организмом хозяина. Регуляция популяцией хозяина.
Команда Ричарда Ленски, микробиолога-эволюциониста, известного своим долгосрочным эволюционным экспериментом, изучила генетические различия (накопленные мутации) между популяциями бактерий, в течение 2000 поколений адаптировавшихся к пяти разным температурным режимам. Несмотря на разнообразие возможных путей адаптации большая часть мутаций оказалась специфичной, то есть они возникали в тех и только тех популяциях, которые развивались при одном и том же или при сходных температурных режимах. Однако большинство этих мутаций позже закрепились и в ходе продолжающегося долгосрочного эволюционного эксперимента. По-видимому, при разной температуре разные мутации оказываются полезнее других (но редко из полезных становятся вредными) - и естественный отбор работает именно с этими различиями в относительной полезности, закрепляя раньше мутации, оказавшиеся в данных условиях более благоприятными.
«Элементы» неоднократно рассказывали о работе лаборатории Ричарда Ленски (Richard Lenski), уже многие десятилетия занимающейся экспериментальным изучением эволюции бактерий. Эти исследования не только позволяют лучше понять закономерности приспособления этих микроорганизмов к среде (которые могут иметь и практическое значение, взять хотя бы устойчивость к антибиотикам), но и дают возможность воочию увидеть нетривиальные эволюционные процессы и их результаты, что важно для фундаментальной науки (см. Ранние этапы адаптации предсказуемы, поздние - случайны , «Элементы», 03.03.2015; В долгосрочном эволюционном эксперименте выявлен отбор на «эволюционную перспективность» , «Элементы», 25.03.2011; Подведены итоги эволюционного эксперимента длиной в 40 000 поколений , «Элементы», 01.11.2009).
Но, кроме широко известного долговременного эволюционного эксперимента (ДЭЭ) (см. E. coli Long-term Experimental Evolution Project), на данный момент длящегося уже 50 000 поколений, команда Ричарда Ленски проводит и более кратковременные исследования, являющиеся «отводками» основного эксперимента.
В новой работе исследователи изучали генетическую основу адаптации кишечной палочки Escherichia coli к разным температурам на протяжении 2000 поколений в нескольких десятках линий. В этой ситуации можно было ожидать как дивергенцию за счет стохастических процессов - возникновения мутаций и генетического дрейфа (см. Genetic drift), - так и параллельную эволюцию , когда разные линии приобретали бы в одной и той же среде сходные адаптации. По крайней мере, для сложно устроенных животных, у которых индивидуальное развитие существенно усложняет путь от генотипа к фенотипу , эти два варианта не обязательно противоречат друг другу: одна и та же адаптация может возникать на разной генетической основе. С этой множественностью допустимых результатов (подчеркнем, что каждый из них оказывается возможным) связан и давно интригующий ученых вопрос о предсказуемости эволюции - вернее, вопрос состоит в том, на каком уровне мы можем предсказывать то, как конкретная популяция приспособится к изменившейся тем или иным образом окружающей среде (см. D. L. Stern, V. Orgogozo, 2009. Is genetic evolution predictable?).
В эксперименте 30 отдельных линий кишечной палочки, взятые из долгосрочного эксперимента, в течение 2000 поколений развивались при пяти разных температурных режимах, по шесть линий на каждый. Часть бактерий выращивали при оптимальных 37°C (напомню, что в норме эти бактерии населяют кишечник теплокровных животных) и «терпимых» 32°C, а часть - при экстремальных для них холоде (20°C) и жаре (42°C), находящихся на нижней и верхней границах зоны толерантности (см. Реакция организма на изменение экологических факторов). Кроме того, еще один режим включал в себя чередование умеренных и экстремальных условий - температура колебалась между 32°C и 42°C. Подробнее схема эксперимента представлена на рис. 2А.
Все популяции бактерий за время эксперимента повысили скорость роста, то есть их приспособленность к среде возросла. Причем обычно эволюционировавшие бактерии оказывались более успешными по сравнению с предковыми не только в той среде, в которой их выращивали, но и на других. Но такое происходило далеко не всегда - например, популяции из 20, 32- и 37-градусных сред при 42°C росли хуже, чем предки. Впрочем, эти результаты были опубликованы уже давно (см. A. F. Bennett, R. E. Lenski, J. E. Mittler, 1992. Evolutionary adaptation to temperature. I. Fitness responses of Escherichia coli to changes in its thermal environment ; J. A. Mongold, A. F. Bennett, R. E. Lenski, 1996. Evolutionary adaptation to temperature. IV. Adaptation of Escherichia coli at a niche boundary). А в свежей работе авторы разбирались в генетической «подноготной» температурных адаптаций. Для этого они полностью отсеквенировали ДНК у представителей каждой популяции из конечного, 2000-го поколения.
Суммарно за все время эксперимента все исследуемые линии накопили 159 мутаций (напомним, что в эксперименте используется популяция E. coli, размножающаяся бесполым путем, поэтому эволюционные процессы изучают по накоплению мутаций) , при этом каждая отдельная линия накопила от двух до восьми мутаций - что интересно, только одна из этих «крайних» популяций росла не при оптимальных 37 градусах, а при «умеренных» 32-х. Но в среднем число мутаций, накопленных 37-градусными бактериями, значимо меньше, чем у всех остальных (но только если рассматривать их всех вместе; при попарном сравнении вариантов эксперимента различия в числе накопленных мутаций оказываются незначимыми - возможно, по причине малого числа популяций в каждом из вариантов опыта). Такие результаты вполне ожидаемы - ведь бактерии росли при оптимальной для них температуре, в которой, к тому же, в течение еще 2000 поколений развивались общие предки экспериментальных линий (рис. 2А). Таким образом, бактерии уже оказывались весьма адаптированными к этим условиям и на них действовал скорее очищающий отбор - отсеивающий мутации, разрушающие уже существующие адаптации, - чем движущий , приводящий к формированию новых приспособлений. Обнаруженные мутации относились к разным типам: это были синонимичные и несинонимичные замены нуклеотидов (см. Мутации: нуклеотидные замены: типы), делеции (утраты участков ДНК) и т. д. Их распределение по типам и отдельным популяциям приведено на рисунке 2В. Три из обнаруженных мутаций могут приводить к гипермутабильности - повышенному темпу мутирования. Такие мутации часто приобретаются в эволюционных экспериментах на бактериях, так как повышают шансы на появление благоприятных мутаций. Однако в данном случае популяции, несущие потенциальные гены-мутаторы , не показали сколько-нибудь значимого превышения числа полученных мутаций над ожидаемым.
Далее ученые оценили сходство мутаций, появившихся в разных популяциях, рассчитав коэффициент сходства Дайса, также известный как коэффициент Сёренсена (S = 2 (X∩Y)/(X + Y), где X и Y - число мутировавших генов в первой и второй сравниваемой группе, а X∩Y - число генов, мутировавших в обеих группах). При этом учитывали только те мутации, которые можно было точно отнести к конкретному гену (например, не учитывались все крупные делеции). Во-первых, коэффициент сходства для популяций внутри каждого варианта опыта оказался существенно больше, чем между разными вариантами: 0,168 против 0,042. При этом статистически незначимой оказалась только разница между линией, культивируемой при 42°C, и культивируемой при переменной температуре (рис. 3А). Во-вторых, удалось выявить специфичные мутации, значимо чаще возникающие (а иногда - и вообще уникальные) для отдельного температурного режима. Всего таковых оказалось четыре - только у переменного температурного режима не оказалось специфичного гена (рис. 3В).
Но, по-видимому, адаптации не сводятся к этим четырем мутациям. Так как часть других мутировавших генов также показывает тенденцию к агрегации в одном из вариантов опыта. Что интересно, все гены, «специфичные» и подозреваемые в этом, относятся к нескольким функциональным группам: они влияют на размер и форму клеток (mrdA , hslU ), регулируют метаболические пути , связанные с переработкой питательных веществ (nadR , iclR ), и меняют активность метаболического фермента (gltB ) (см. рис. 3В). Также среди «холодных» популяций есть тенденция к мутированию генов, связанных с транскрипцией и трансляцией . Таким образом, перед нами типичный пример комплексной адаптации, задействующей разные механизмы - и потому потенциально многовариантной. К счастью, современные молекулярно-генетичские методы позволяют вычленять отдельные гены и признаки, участвующие в приспособлении к тому или иному фактору среды, хотя еще несколько десятков лет назад едва ли не все эволюционисты считали, что распутать этот клубок невозможно и точные механизмы таких сложных адаптаций непознаваемы. Действительно, тогда можно было только сказать что-то вроде: «в ответе на отбор участвует множество генов, в каждой популяции - своих». Сейчас же мы можем сказать, какие именно это гены. И оказывается, что по крайней мере в некоторых случаях этот набор участвующих в адаптации генов оказывается воспроизводимым.
Интересные результаты принесло сравнение описываемого эксперимента с продолжающимся долговременным эволюционным экспериментом. Оказалось, что бактерии из ДЭЭ со временем накапливают мутации в тех же генах, что и адаптирующиеся к разным температурам! С одной стороны, им нужно адаптироваться не только к температуре - например, в среде обоих экспериментов не хватает глюкозы. С другой - где же тут специфичность адаптаций? Да, линия, культивируемая при 37°С, показывает наибольшее сходство с ДЭЭ, протекающем при той же температуре, - но ведь и остальные приобретают мутации, которые впоследствии оказываются полезными и при других температурах (рис. 4).
Авторы объясняют этот парадокс тем, что разные температурные режимы меняют относительную ценность разных мутаций, но редко делают полезную мутацию вредной - то есть, например, мутация, закрепившаяся в «холодных» линиях, в условиях низких температур оказывается более благоприятной, чем в оптимальных условиях, что повышает ее шанс на более раннее закрепление. В пользу этого говорит и прямая оценка адаптивной ценности отдельных мутаций. Исследователи вводили их в геном предковых бактерий и смотрели, как эти модифицированные организмы чувствуют себя при разных температурах. Ни для одной из мутаций не обнаружилось значимых различий по положительному эффекту на скорость роста при разных температурных режимах, хотя попарные сравнения выявили слабую тенденцию к тому, чтобы мутация была более благоприятной в том варианте опыта, где она возникла. Это подтверждает интерпретацию авторов: со сменой температуры все закрепившиеся мутации остаются благоприятными, но их относительная полезность меняется и естественный отбор работает именно с ними: оказавшиеся более благоприятными в данных условиях мутации закрепляются раньше.
Это исследование позволяет еще раз подчеркнуть, что эволюция может идти за счет вроде бы слабых, малозаметных различий. Не наличие эффективных и неэффективных путей, но небольшие различия в эффективности тех или иных приспособлений при разных температурах меняют вероятности их закрепления естественным отбором, что приводит к разным путям эволюции.
В конце статьи авторы подчеркивают, что, учитывая широкий диапазон возможностей для адаптации, их результаты показывают неожиданно высокую предсказуемость эволюции. И предполагают, что наличие таких специфических следов адаптации к конкретным условиям среды может помочь, например, в расшифровке эволюционной истории патогенных микробов, отслеживая историю перехода между разными видами, или даже индивидами хозяев (что поможет отследить динамику заболеваний), а также в судебной микробиологии, помогая выяснять обстоятельства гибели или историю перемещений отдельных предметов (см. R. E. Lenski, P. Keim, 2005.
Биологическая адаптация (от лат. adaptatio - приспособление) - приспособление микроорганизма к внешним условиям в процессе эволюции, включая морфофизиологическую и поведенческую составляющие. Адаптация может обеспечивать выживаемость в условиях конкретного местообитания, устойчивость к воздействию факторов абиотического и биологического характера, а также успех в конкуренции с другими видами, популяциями, особями. Каждый вид имеет собственную способность к адаптации, ограниченную физиологией (индивидуальная адаптация).
Дезадаптация – какое-либо нарушение адаптации, приспособление организма к постоянно меняющимся условиям внешней или внутренней среды. Состояние динамического несоответствия между живым организмом и внешней средой, приводящее к нарушению физиологического функционирования, изменению форм поведения, развитию патологических процессов Полное несоответствие между организмом и внешними условиями его существования несовместимо с жизнедеятельностью. Степень дезадаптации характеризуется уровнем дезорганизации функциональных систем организма. В зависимости от характера функционирования, выделяют две формы дезадаптации: – непатологическая: поддержание гомеостаза возможно при режиме усиленного, но "нормального" физиологического функционирования; – патологическая: поддержание гомеостаза возможно только при переходе к патологическому функционированию.
Адаптации видов в рамках одного биоценоза зачастую тесно связаны друг с другом. Если адаптационный процесс у какого-либо вида не находится в равновесном состоянии, то эволюционировать может весь биоценоз (иногда - с негативными последствиями) даже в стабильных условиях окружающей среды.
Главное содержание адаптации, по мнению Т. Пилат, - это внутренние процессы в системе, которые обеспечивают сохранение её внешних функций по отношению к среде. Если структура системы обеспечивает ей нормальное функционирование в данных условиях среды, то такую систему следует считать адаптированной к этим условиям. На этой стадии устанавливается динамическое равновесие.
Примеры адаптации: у пресноводных простейших осмотическая концентрация протоплазмы выше, чем концентрация, окружающей воды. При поглощении воды в ней происходит постоянное опреснение. Нарушающееся при этом осмотическое равновесие регулируется деятельностью сократительной вакуоли, которая удаляет из тела избыточную воду. Некоторые простейшие способны, однако, приспособляться к существованию в более соленой и даже морской воде. При этом деятельность сократительной вакуоли у них замедляется и может даже совершенно прекратиться, так как в этих условиях выведение из организма воды повело бы к повышению в протоплазме относительной концентрации ионов и, в связи с этим, к нарушению в ней осмотического равновесия. Таким образом, в этом случае, механизм адаптации сводится к непосредственной физико-химической реакции протоплазмы. В других случаях механизм адаптации представляется более сложным и не всегда может быть сразу разложен на элементарные факторы. Таковы, например, адаптация животных к температурным условиям (удлинение шерсти млекопитающих под влиянием холода), к явлениям лучистой энергии (фототропизм растений); изменение окраски кожи холоднокровных, благодаря реакции пигментных клеток; сезонный диморфизм окраски птиц и млекопитающих; изменение их окраски в зависимости от климато-географических условий и т. п. Однако, и здесь механизм адаптации может быть, в конечном счете, сведен к физико-химическим реакциям протоплазмы. Явления адаптации тесно связаны с эволюцией микроорганизмов и представляют собою один из наиболее существенных факторов акклиматизации, борьбы за существование и мимикрии.
Адаптация микроорганизмов, аккомодация микроорганизмов, приспособление их к окружающей среде. Их строение, физиологические свойства и химимический состав находятся в зависимости как от наследственных свойств данного вида, так и от влияний окружающей среды. Последние заставляют микроорганизм изменяться. Изменения эти еще недавно считались случайными и, по учению Кона (Conn), мало существенными для основных черт микроорганизма, которые признавались незыблемыми. Однако, с течением времени сначала робко, а затем все решительнее, было выдвинуто учение об изменчивости микроорганизмов, как биологическом факторе, и в настоящее время изменения микроорганизмов уже не считаются только случайными, а признаются и более глубокими. Характер изменчивости микроорганизма находится в зависимости от двух факторов: от индивидуальной видовой стойкости данного микроорганизма и от глубины, размаха и силы воздействия окружающей среды. Некоторые виды микроорганизмов, как кислотоупорная группа, дифтерийная и грибковая формы, меньше изменяются и хуже приспособляются, в то время как кишечно-тифозная, капсулярная, кокковая, анаэробная группы легче подвергаются изменениям. Приспособляемость микроорганизмов прежде всего сказывается в их отношении к кислороду и окружающей температуре. Известно, что анаэробы могут быть приучены как к свободному кислороду, так и обратно. То же нужно сказать об отношении к окружающей температуре, а также к реакции среды, к действию света и химическому составу питательного материала. Одно условие должно быть соблюдено для выявления этой адаптации: постепенное воздействие новых факторов. Чем медленнее и постепеннее действуют новые условия, тем легче и совершеннее приспособляется микроорганизм. Это приспособление идет в различных направлениях. Окружающие условия заставляют микроорганизм становиться менее требовательным в своих физиологических функциях, ограничивать их до минимума и переходить в стадию анабиоза («скрытый микробизм»), для чего у него образуются споры, и он окружается непроницаемыми слизистыми, известковыми и соединительнотканными капсулами (кокки, туб. палочки и т. п.); или же микроорганизмы претерпевают морфологические изменения, теряя целые органы и части, особо чувствительные к обычным условиям (например, трипанозомы, приучаясь к мышьяку, теряют блефаробласты (Вербицкий)), и, таким образом, получаются новые расы микроорганизмов. Образование новых рас с новыми свойствами происходит особенно легко, когда микроорганизм встречается с новыми химическими веществами в организме, в котором он привык свободно размножаться. Когда в такой среде появляются вредные вещества, часть микроорганизмов погибает, а наиболее стойкие индивидуумы выживают и дают так называемые «стойкие» или «упорные» расы (Enrlich). Такая стойкость доказана в отношении различных химических соединений и алкалоидов (мышьяк, спирт, хинин).- Приспособляемость микроорганизмов может идти и в противоположном направлении - в сторону усиления их жизнеспособности и приобретения ими большей активности. Так, мало вирулентный микроорганизм под влиянием ослабления организма начинает быстро размножаться и вырабатывать токсины, которых у него раньше не было или было мало. Примером здесь могут служить многочисленные случаи так называемые эндогенных инфекций, когда пневмококк под влиянием простуды вызывает пневмонию или Bact. coli под влиянием погрешности в диете вызывает дизен-терийноподобное заболевание. Это «активирование» микроорганизма есть не что иное, как приспособление его к новым условиям. Явления приспособления особенно хорошо изучены и многочисленны там, где микроорганизм встречается с иммунным организмом или иммунными средами. Кроме указанных выше капсул, служащих микробу защитным слоем от внешней среды, у микроорганизма начинают вырабатываться аггрессины, которые делают его мало доступным для фагоцитов. Приспособляемость микроорганизмов идет так далеко, что они могут делаться стойкими даже по отношению к иммунным сывороткам. Борде (Bordet) еще в 1895 г. показал, как холерный вибрион может быть приучен к бактериолитической сыворотке. Рядом авторов доказана возможность приучить аглютинирующихся микроорганизмов к тому, что они перестают аглютинироваться. И наоборот, неаглютинабильные микроорганизмы могут быть превращены в аглютинирующихся, например, путем проведения через организм животных и даже при простых пересевах из среды на среду. Перестраивая свои морфологические и физиологические черты, микроорганизмы в зависимости от почвы, на которой они живут, и в зависимости от других микроорганизмов, рядом с ним размножающихся, может приобрести черты, присущие соседу, и превратиться в так называемый «парамикроба». Такой микроорганизм, как это доказал Розенау (Rosenow), может приобрести новые свойства, полученные им от сожительства с патогенным микроорганизмом, и сохранять их довольно долго по наследству. Так, например, стрептококк, выделенный при менингитах, вызванных диплококком Weichselbaum’a, приобретает способность давать менингит. Получается как бы подражание другому возбудителю. Это подражание выражается или в способности вызывать такое же заболевание или в приобретении новых антигенных свойств. Так, протей, живущий в организме сыпнотифозного больного, начинает аглютинироваться сывороткой больного, хотя он и не является возбудителем болезни. Из всех приведенных фактов ясно, какое большое значение имеют явления адаптации микроорганизмов для патологии и эпидемиологии.
Эволюция бактерий и ее медицинское значение. Микроорганизмы на Земле возникли примерно за три миллиарда лет до появления человека. В 1822 году Э.Дарвин предложил теорию эволюции, а через 100 лет русский биохимик А.Опарин (1920 г.) – теорию возникновения биологической жизни. В этой системе бактериям принадлежит весьма важное место. Первые, окруженные мембраной самореплицирующиеся формы биологической жизни (протобионты) были неспособны к фотосинтезу и получали энергию путем осуществления простых, одностадийных абиогенных окислительных реакций. Это продолжалось около 1.0 млрд. лет. Энергия (электрохимическая, термальная, фотохимическая), образующаяся в этих реакциях, сохранялась в определенных молекулах и использовалась для осуществления примитивных процессов. Формирование первичных молекул и реакций положило начало обменным процессам-анаболизму и катаболизму. Переход от протоклетки к прокариотной клетке произошел в промежутке 2.5-3 млрд. лет назад. В атмосфере планеты не было кислорода и первичные прокариоты были анаэробами. Аутотрофный путь фиксации СО 2 явился основой первичной продуктивности на планете. Смена восстановительной атмосферы на кислородную произошла между средним и поздним докембрием (2,8 млрд. лет назад). Для сравнения содержание кислорода в атмосфере планеты 800 млн. лет назад составляло около 1%, 400 млн. лет – уже было 10%, а в настоящее время – 21%. По мере изменения состава атмосферы стали формироваться факультативные фототрофные и гетеротрофные анаэробы, позднее возникли аэробные бактерии .
Бактерии явились не только первичными накопителями генов, но объектом их эволюционного усовершенствования. Скорость эволюции – это количество мутаций на 100 аминокислот молекулы определенного белка в течение 100 млн. лет . Она широко варьирует. На этом построена концепция молекулярных часов, декларирующая, что мутации постепенно аккумулируются в геноме и линейно временному периоду эволюции формируют новый сиквенс для дальнейшей дивергенции вида. Диаграмма, представленная на рис.3. позволяет отобразить эволюцию определенных групп бактерий и примерно установить эволюционное время, когда тот или иной вид (род) дивергировал от общего предка .
Скорость эволюции постоянна и зависит от многих факторов – скорости метаболических процессов, времени генерации, потоков движения информации и селективного давления. Например, дивергенция рода Salmonella и рода Escherichia coli от общего предка произошла примерно 100-140 млн лет назад. Геномы бактерий эволюционировали на протяжении более 50 миллиардов генераций аккумулируя мутации и приобретая новую генетическую информацию посредством горизонтального переноса генов без существенной перестройки предковых генов. В течение года геном сальмонелл приобретал чужеродной генетической информации примерно 16 кб/млн. лет, а кишечной палочки – 22 кб/млн.лет. В настоящее время их геномы отличаются на 25% . Значительная часть генома приобретена путем горизонтального переноса. В целом, геном бактерий варьирует по размерам от 0.6 до 9.4 Мб информации (в среднем от 3 до 5 Мб). Некоторые бактерии имеют две хромосомы (Leptospira interrogans serovar icterohemorrahgiae, Brucellae melitensis) . Прогрессивная эволюция бактерий происходила в нескольких взаимосвязанных направлениях – метаболическом, морфологическом (структурно-молекулярном) и экологическом. В природе имеется огромное разнообразие микроорганизмов из которого в настоящее время известно не более 5-7% их, а культивируемые в искусственных условиях бактерии составляют около 1%. Это означает, что мы еще только начинаем узнавать мир микробов.
Стратегии секвенирования генома. Каждая пара оснований генома является одним битом информации. Например, геном Haemophilus influenzae содержит 1 830 137, а геном Escherichia coli – 4 639 221 бит информации. Сравнительные аспекты секвенирования геномов бактерий позволяют определить наличие общих генов, регуляторных механизмов, установить эволюционные внутри и межвидовые связи и являются основой структурной и эволюционной геномики. Математическим анализом геномов микроорганизмов занимается новая наука – биоинформатика. Предметом исследований являются сиквенсы фрагментов или полных геномов бактерий с помощью разрабатываемых компьютерных программ и баз данных информации о нуклеиновых кислотах и белках .
На основе анализа строения геномов (секвенирования) сформировано 36-40 крупных таксонов (отделов). Члены каждого из них имеют общего предка, который на определенном этапе дивергировал от другого таксона-предшественника. Некоторые из отделов включают большее число видов известных бактерий, чем другие. Обычно это относится к тем из них, которые хорошо культивируются в лабораторных условиях. Наибольшее число видов бактерий (от 40 до 80%) описано среди таксонов протеобактерий, актинобактерий, грамположительных бактерий с низким содержанием Г+Ц. Вместе с тем в некоторых отделах культивируемые представители бактерий неизвестны. Следует отметить, что из 36-40 отделов царства Bacteria только представители 7 крупных таксонов способны вызывать заболевания у человека. Специализация и адаптация этих бактерий к организму животных привело к образованию блоков генов, контролирующих факторы патогенности (островки патогенности). Они могут локализоваться в хромосоме, плазмидах и, возможно, в фагах бактерий. Установление направления и порядка эволюции микроорганизмов на основе изменчивости их геномов является перспективным направлением молекулярной эпидемиологии .
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки |  | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.
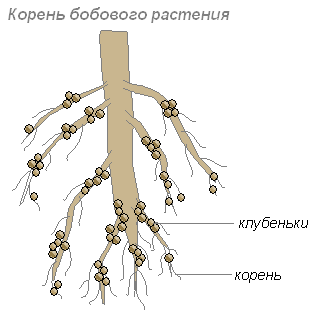
Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.
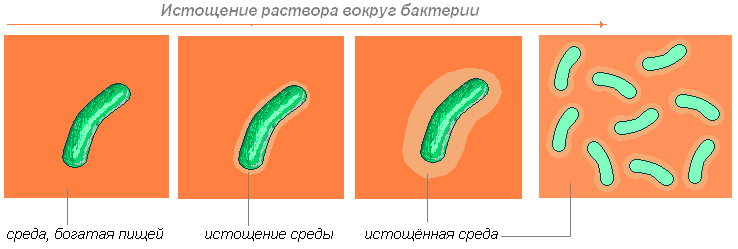
Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.